

 A new skull of the fossil porpoise Numataphocoena yamashitai (Cetacea: Phocoenidae) from the upper part of the Horokaoshirarika Formation (lower Pliocene), Numata Town, Hokkaido, Japan, and its phylogenetic position
A new skull of the fossil porpoise Numataphocoena yamashitai (Cetacea: Phocoenidae) from the upper part of the Horokaoshirarika Formation (lower Pliocene), Numata Town, Hokkaido, Japan, and its phylogenetic position
Article number: 19.3.49A
https://doi.org/10.26879/663
Copyright Society for Vertebrate Paleontology, November 2016
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 March 2016. Acceptance: ??
{flike id=1663}
ABSTRACT
An early Pliocene porpoise, Numataphocoena yamashitai from Hokkaido, Japan, is known from the holotype, a fairly well-preserved skeleton with an incomplete skull and a referred earbone. A new skull referred to Numataphocoena yamashitai found from almost the same locality as the holotype is interesting because it expands knowledge of skull morphology and improves the diagnosis of this taxon. Numataphocoena yamashitai differs from other phocoenids in having the characteristic feature in the maxilla associated with the posterior dorsal infraorbital foramen, narrower and sharper anterior part of the internal acoustic meatus, and a robust anterior process of the periotic. A new cladistic analysis places Numataphocoena yamashitai adjacent to Haborophocoena toyoshimai and Haborophocoena minutus, among a clade of early branching phocoenids, all of which are chronologically and geographically close to each other. The new skull is probably a younger individual because it is about 80% the size of that of the holotype and it shows closed but unfused sutures. Our description of this specimen helps to understand the intraspecies variation of the extinct species Numataphocoena yamashitai.
Yoshihiro Tanaka. Numata Fossil Museum, 2-7-49, Minami 1, Numata Town, Hokkaido, 078-2225 Japan, yoshihiro.tanaka@otago.ac.nz and Hokkaido University Museum, Kita 10, Nishi 8, Kita-ku, Sapporo, Hokkaido 060-0810 Japan
Hiroto Ichishima. Fukui Prefectural Dinosaur Museum, Terao 51-11, Muroko, Katsuyama, Fukui 911-8601, Japan, hiroto.ichishima@dinosaur.pref.fukui.jp
Key words: skull; Phocoenidae; phylogeny; maxillary terrace; ontogeny; intraspecies variation
Final citation: Tanaka, Yoshihiro and Ichishima, Hiroto. 2016. A new skull of the fossil porpoise Numataphocoena yamashitai (Cetacea: Phocoenidae) from the upper part of the Horokaoshirarika Formation (lower Pliocene), Numata Town, Hokkaido, Japan, and its phylogenetic position. Palaeontologia Electronica 19.3.49A: 1-28. https://doi.org/10.26879/663
palaeo-electronica.org/content/2016/1663-a-new-skull-of-numataphocoena
Copyright: © December 2016 Society of Vertebrate Paleontology. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. creativecommons.org/licenses/by/4.0/
INTRODUCTION
Since 2000, fossil phocoenid study has been progressed dramatically (Ichishima and Kimura, 2000, 2005, 2009, 2013; Fajardo-Mellor et al., 2006; Lambert, 2008; Murakami et al., 2012a, 2012b, 2015; Racicot et al., 2014; Colpaert et al., 2015). Fossil phocoenids have long been known only from the eastern and southern Pacific since the study of Salumiphocaena stocktoni (Barnes, 1985a) from the late Miocene of California by Wilson (1973), a series of Piscolithax spp reported from Peru and Mexico (de Muizon, 1983; Barnes, 1984a), and an isolated periotic described from the late Miocene of New Zealand (Fordyce, 1989). Recently, a phocoenid with a unique feeding apparatus was reported from California (Boessenecker, 2013; Racicot et al., 2014). Now, a few specimens have been known from the Atlantic (Lambert, 2008; Colpaert et al., 2015).
Tomida and Kohno (1992) described a skull from Wakkanai, Hokkaido, Japan, the first fossil record of Phocoenidae from the western North Pacific. After that, six species of fossil phocoenid have been published from Hokkaido (Ichishima and Kimura, 2000, 2005, 2009, 2013; Murakami et al., 2012a, 2012b, 2015).
Numataphocoena yamashitai from the early Pliocene of Numata Town, Hokkaido, Japan (Ichishima and Kimura, 2000) is a fairly well-preserved specimen represented by the partial skull, ear bones and postcrania. But it was too premature to perform the phylogenetic analysis for the holotype of Numataphocoena yamashitai at the time of description because of the incompleteness of the skull, which hampered a direct comparison with most other fossil phocoenids represented by the skull but lacking postcrania. A new skull, along with a referred periotic collected from the same locality (Tanaka, 2016), of Numataphocoena yamashitai allows us to determine the phylogenetic placement of the species.
Abbreviation. NFL - Numata Fossil Museum, Hokkaido, Japan.
MATERIAL AND METHODS
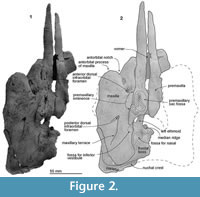
 Numata Fossil Museum specimen NFL 2074, a skull; the premaxillae, left posterior maxilla, left frontal, left lacrimojugal, left palatine and left parietal. The specimen was originally prepared by Mr. Shigeru Yamashita using formic acid. Additional preparation was done by the first author under microscope with a fine air chisel.
Numata Fossil Museum specimen NFL 2074, a skull; the premaxillae, left posterior maxilla, left frontal, left lacrimojugal, left palatine and left parietal. The specimen was originally prepared by Mr. Shigeru Yamashita using formic acid. Additional preparation was done by the first author under microscope with a fine air chisel.
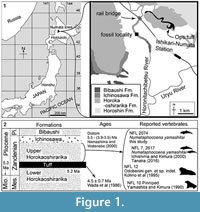
NFL 2074 was collected from the riverbed of the Horonitachibetsu River, early Pliocene, upper part of the Horokaoshirarika Formation, at 4th Ebishima district, Numata Town, Hokkaido, Japan by Dr. Rei Nakashima in 1995. The site is about 3 km northwest of the Ishikari-Numata Station of the Japan Railroad (Figure 1).
Geological setting. In Numata Town, Hokkaido, the Miocene-Pliocene sediments, the upper and lower parts of the Horokaoshirarika Formation are about 200 m and 500 m in thickness, respectively (Watanabe and Yoshida, 1995) (Figure 1). A thin tuff, so-called Ops, separates the Horokaoshirarika Formation into the upper and lower parts (Kobayashi et al., 1969). This tuff layer is dated as 4.5 ± 0.7 Ma, based on fission track method (Wada et al., 1986). In the lower stream of the Horonitachibetsu River, the Ichinosawa and Bibaushi Formations overlie the Horokaoshirarika Formation. The Horoshin Formation is exposed about 2 km upstream from this locality. NFL 2074 was collected from the upper part of the Horokaoshirarika Formation. The upper part of the Horokaoshirarika Formation yielded other marine vertebrate remains: the holotype (NFL 7) and an isolated periotic (NFL 2617) of Numataphocoena yamashitai, a tusk of Odobenini (NFL 12) and a pinniped skeleton (NFL 10) have been reported (Yamashita and Kimura, 1990; Kohno et al., 1995; Ichishima and Kimura, 2000; Tanaka, 2016). Lithology of the upper part of the Horokaoshirarika Formation is muddy to sandy sediments, with shell clusters and bioturbation (Nakashima and Majima, 2000). Paleoenvironment of the upper part of the Horokaoshirarika Formation was inner shelf (Nakashima and Majima, 2000). As it is mentioned above, Wada et al. (1986) reported the tuff age as 4.5 ± 0.7 Ma based on the fission track dating. Based on diatom biostratigraphy, the age of the upper part of the Horokaoshirarika Formation is 5.5 to 3.5 Ma, corresponding to the Thalassiosira oestrupii zone (Nakashima and Watanabe, 2000). The age of the upper part of the Horokaoshirarika Formation is about 4.5 to 3.5 Ma, the early Pliocene.
SYSTEMATIC PALEONTOLOGY
Order CETACEA Brisson, 1762
Unranked taxon NEOCETI Fordyce and de Muizon, 2001
Suborder ODONTOCETI Flower, 1867
Superfamily DELPHINOIDEA (Gray, 1821) Flower, 1867
Family PHOCOENIDAE Gray, 1825, sensu Burmeister, 1885
Numataphocoena yamashitai Ichishima and Kimura, 2000
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Table 1
 Emended diagnosis. Numataphocoena yamashitai differs from other phocoenids in having a raised area along an extended sulcus from the medial border of the posterior dorsal infraorbital foramen on the dorsal surface of the maxilla, the maxillary terrace (new term, see Discussion); narrower and sharper anterior part of the internal acoustic meatus; and a robust anterior process of the periotic. Numataphocoena yamashitai differs from later branching phocoenids (such as Lomacetus, Piscolithax spp, extant species) in absence of the maxillary crest (Character 28), and wide premaxillae against the maxillae at the level of the postorbital process (Character 35).
Emended diagnosis. Numataphocoena yamashitai differs from other phocoenids in having a raised area along an extended sulcus from the medial border of the posterior dorsal infraorbital foramen on the dorsal surface of the maxilla, the maxillary terrace (new term, see Discussion); narrower and sharper anterior part of the internal acoustic meatus; and a robust anterior process of the periotic. Numataphocoena yamashitai differs from later branching phocoenids (such as Lomacetus, Piscolithax spp, extant species) in absence of the maxillary crest (Character 28), and wide premaxillae against the maxillae at the level of the postorbital process (Character 35).
Description
Morphological terminology for the skull follows Mead and Fordyce (2009).
Ontogenetic age. The skull sutures are mostly closed but distinct in NFL 2074. Compared with the holotype of Numataphocoena yamashitai (NFL 7), which is a physically and sexually matured individual (Ichishima and Kimura, 2000), NFL 2074 is around 20% smaller in size, based on the length of the maxilla (the distance between the anterior tip of the antorbital process of the maxilla to the posterior end of the ascending process. NFL 2074: 145 mm; NFL 7: 188 mm) (Table 1). Ichishima and Kimura (2013) reported the referred specimen of Haborophocoena toyoshimai as a young individual on the basis of the incompletely fused supra/exoccipital suture, which is around 16% smaller skull size than the physically matured holotype of Haborophocoena toyoshimai. NFL 2074 is, therefore, most likely younger than the holotype.
Premaxilla. The distance between the tip of the premaxilla as preserved and the base of the rostrum at the level of the antorbital notch is 106 mm. Each premaxilla is flat anterior to the level of the antorbital notch, and posteriorly it rises dorsally as the premaxillary eminence, which projects dorsolaterally with a weak depression on the dorsal face (the premaxillary sac fossa). In dorsal view, the posterior end of the premaxillae is widest (27.0 mm on the left) at the level of the nares. A rounded end of the nasal process stops at the level of the posterior margin of the bony nares. Ventrally, the anterior part shows sutures with the lost maxilla laterally, and with the vomer medially.
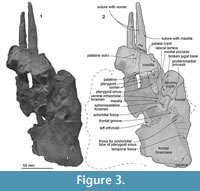 Maxilla. The preserved cranial part of the left maxilla is dorsoventrally thin and rises gradually posterodorsally at the level of the bony nares. The base of the rostrum is wide and has a distinct antorbital notch. On the ventral face, the maxilla forms a part of the palate, which is flat anteriorly and has a weak palatal crest posteriorly. Two shallow palatine sulci run anteroposteriorly on the right maxilla. A rounded antorbital process projects anteriorly and forms a sharp and deep antorbital notch medially. Medial to the antorbital process, there is an anterior dorsal infraorbital foramen.
Maxilla. The preserved cranial part of the left maxilla is dorsoventrally thin and rises gradually posterodorsally at the level of the bony nares. The base of the rostrum is wide and has a distinct antorbital notch. On the ventral face, the maxilla forms a part of the palate, which is flat anteriorly and has a weak palatal crest posteriorly. Two shallow palatine sulci run anteroposteriorly on the right maxilla. A rounded antorbital process projects anteriorly and forms a sharp and deep antorbital notch medially. Medial to the antorbital process, there is an anterior dorsal infraorbital foramen.
In dorsal view, the maxilla covers most of the frontal. The posteromedial surface of the maxilla is steep at the level of the bony nares. The posterior dorsal infraorbital foramen opens into a groove, which continues posterolaterally to an area, the maxillary terrace (new term, see Discussion), whose posterior margin reaches the lateral edge of the skull roofing over the temporal fossa. The maxillary intrusion (sensu Arnold and Heinsohn, 1996), a dorsal exposure of the maxilla medial to the premaxilla and anterior to the bony nares is uncertain. An incipient fossa for the inferior vestibule (Mead, 1975) is just posterior to the nasal process of the premaxilla and lateral to the bony nares. The fossa for the inferior vestibule is circular and much shallower than that of modern phocoenids, which have a small expansion medially.
In ventral view, just posteromedial to the lacrimojugal is the antorbital fossa, which includes the ventral infraorbital foramen anteriorly and the sphenopalatine foramen posteriorly.
Palatine. Ventrally, the left palatine shows a smooth anterior wall of the pterygoid sinus fossa, which is a dorsoventrally long elliptical fossa.
Pterygoid. The left anterior fragment of the pterygoid might be on the skull, just posterior to the palatine, but the suture is not clear.
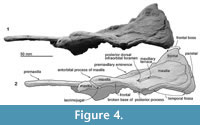 Ethmoid. The ethmoid is used in the sense of Mead and Fordyce (2009), but note that Ichishima (2011) suggested that the mesethmoid is probably absent in Odontoceti. The structure of the preserved left ethmoid is unclear in the specimen because of erosion and damage. A thin imperforated cribriform plate forms the posterior wall of the nasal passage. The cribriform plate rises to the fossa for nasals on the frontal. The dorsal end shows a partially damaged osseous nasal septum.
Ethmoid. The ethmoid is used in the sense of Mead and Fordyce (2009), but note that Ichishima (2011) suggested that the mesethmoid is probably absent in Odontoceti. The structure of the preserved left ethmoid is unclear in the specimen because of erosion and damage. A thin imperforated cribriform plate forms the posterior wall of the nasal passage. The cribriform plate rises to the fossa for nasals on the frontal. The dorsal end shows a partially damaged osseous nasal septum.
Vomer. An anterior broken section of the vomer can be seen in the mesorostral groove, which is V-shaped in anterior view. The most posterior part of the vomer has been worn away.
Sphenoid. Posteromedial to the orbital rim, there is a shallow and mediolaterally long groove, which might be the frontal groove of the sphenoid. In general, the frontal groove runs from a combined large foramen of the orbital fissure and optic canal, but the medial part of the frontal groove is broken away on NFL 2074.
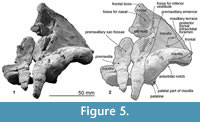 Frontal. The frontal contributes to the ventral surface of the orbit and the frontal boss at the vertex. The frontal boss is smooth and was originally bounded by the nasals and maxillae. The frontal forms the anterodorsal wall of the braincase and ventrally exposes around a half of the shallow and long orbit. Posterior to the frontal groove, there is a shallow fossa for the postorbital lobe of the pterygoid sinus, just anterior to the temporal fossa. The nasals are not preserved, and their articular surfaces on the frontal are shallow. The fossa for the nasal is anteroposteriorly longer than wide (around 2.0 cm long, 1.5 cm wide). The posteromedial corner of the nasal might be positioned more anterior than the posterolateral corner.
Frontal. The frontal contributes to the ventral surface of the orbit and the frontal boss at the vertex. The frontal boss is smooth and was originally bounded by the nasals and maxillae. The frontal forms the anterodorsal wall of the braincase and ventrally exposes around a half of the shallow and long orbit. Posterior to the frontal groove, there is a shallow fossa for the postorbital lobe of the pterygoid sinus, just anterior to the temporal fossa. The nasals are not preserved, and their articular surfaces on the frontal are shallow. The fossa for the nasal is anteroposteriorly longer than wide (around 2.0 cm long, 1.5 cm wide). The posteromedial corner of the nasal might be positioned more anterior than the posterolateral corner.
Lacrimojugal. The lacrimojugal is squared in ventral view and thin in lateral view. Its ventral surface is eroded. Anteromedially, there is a broken base of the lacrimojugal (8 mm diameter). The anterior border of the lacrimojugal forms the most anterior part of the antorbital process. The tubacular posteromedial process is located posterolaterally. The medial process locates medial and just a bit anterior to the posteromedial process, and is covered by the maxilla medially.
Parietal. The preserved parietal is exposed as an anteroposteriorly narrow band dorsally, just posterior to the frontal. The parietal forms the dorsal part of the temporal fossa. The parietal/frontal suture is unclear at the posterolateral part of the skull. The nuchal crest of NFL 2074 is formed by the parietal dorsolaterally and might be formed by the frontal medially. The supraoccipital is lost in NFL 2074, which might also form the nuchal crest.
RESULTS
Phylogenetic Analysis
 The phylogenetic position of Numataphocoena yamashitai is analyzed using a new data matrix, which is modified from Tanaka and Fordyce (2015), for understanding relationships of the Phocoenidae (see Appendix 1, Appendix 2, Appendix 3, Appendix 4, Appendix 5). In the matrix, three specimens of Numataphocoena yamashitai (NFL 7, the holotype; NFL 2074, the skull of our study; and NFL 2617, a periotic) are combined. There are no contradictive codings among these specimens. Semirostrum ceruttii (Racicot et al., 2014) and Borabocetus gigaseorum (Colpaert et al., 2015) are added in the matrix. Some changes of character coding and deletions of characters from the original matrix are listed in Appendix 4.
The phylogenetic position of Numataphocoena yamashitai is analyzed using a new data matrix, which is modified from Tanaka and Fordyce (2015), for understanding relationships of the Phocoenidae (see Appendix 1, Appendix 2, Appendix 3, Appendix 4, Appendix 5). In the matrix, three specimens of Numataphocoena yamashitai (NFL 7, the holotype; NFL 2074, the skull of our study; and NFL 2617, a periotic) are combined. There are no contradictive codings among these specimens. Semirostrum ceruttii (Racicot et al., 2014) and Borabocetus gigaseorum (Colpaert et al., 2015) are added in the matrix. Some changes of character coding and deletions of characters from the original matrix are listed in Appendix 4.
The matrix includes 22 taxa (Kentriodon pernix as an out group) and 122 characters cited and/or modified from previous studies (Flower, 1867, 1884; Allen, 1923; Miller, 1923; Fraser and Purves, 1960; Noble and Fraser, 1971; Kasuya, 1973; De Smet, 1977; Whitmore and Sanders, 1977; Barnes, 1984a, 1984b, 1985a, 1985b, 1990; de Muizon, 1984, 1985, 1987, 1988, 1991, 1994; Heyning, 1989, 1997; Marsh et al., 1989; Rommel, 1990; Curry, 1992; Fordyce, 1994, 2002; Arnold and Heinsohn, 1996; Luo and Marsh, 1996; Messenger and McGuire, 1998; Sanders and Barnes, 2002; Geisler and Sanders, 2003; Bianucci, 2005; Lambert, 2005, 2008; Fajardo-Mellor et al., 2006; Aguirre-Fernández et al., 2009; Mead and Fordyce, 2009; Geisler et al., 2011, 2012; Murakami et al., 2012b, 2012a, 2014; Tanaka and Fordyce, 2014, 2015). Twelve characters from Colpaert et al. (2015) and two novel characters are added to the matrix of Tanaka and Fordyce (2015).
Character data and tree data were managed using Mesquite 2.75 (Maddison and Maddison, 2011). An analysis was performed with TNT 1.1 (Goloboff et al., 2008). All characters were treated as unweighted and unordered. The analysis used New Technology Search with the setting: recover minimum length trees = 1000 times.
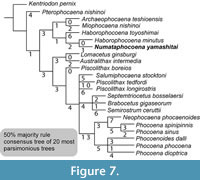 The unweighted and unordered phylogenetic analysis shows 20 shortest trees of 238 steps. A tree file is provided as Appendix 5. The 50% majority rule consensus tree (Figure 7) shows similar topology as for the result of the analysis 2 in Tanaka and Fordyce (2015) such that Pterophocaena nishinoi is placed as the earliest diverging phocoenid, all phocoenids from Hokkaido, Japan are placed as a clade, Lomacetus and Australithax are monophyletic, and extant species are monophyletic. Differences from Tanaka and Fordyce (2015) included Archaeophocaena and Miophocaena monophyly (sister taxon relationship), having an unresolved polytomy of Septemtoriocetus + Brabocetus + Semirostrum + Piscolithax longirostris + Piscolithax tedfordi + (Lomacetus + Australithax) + an extant clade, but having resolved extant clades.
The unweighted and unordered phylogenetic analysis shows 20 shortest trees of 238 steps. A tree file is provided as Appendix 5. The 50% majority rule consensus tree (Figure 7) shows similar topology as for the result of the analysis 2 in Tanaka and Fordyce (2015) such that Pterophocaena nishinoi is placed as the earliest diverging phocoenid, all phocoenids from Hokkaido, Japan are placed as a clade, Lomacetus and Australithax are monophyletic, and extant species are monophyletic. Differences from Tanaka and Fordyce (2015) included Archaeophocaena and Miophocaena monophyly (sister taxon relationship), having an unresolved polytomy of Septemtoriocetus + Brabocetus + Semirostrum + Piscolithax longirostris + Piscolithax tedfordi + (Lomacetus + Australithax) + an extant clade, but having resolved extant clades.
The result of the phylogenetic analysis in our study is compared with previous studies (e.g., Fajardo-Mellor et al., 2006; Lambert, 2008; Murakami et al., 2012a, 2012b, 2014, 2015; Racicot et al., 2014; Colpaert et al., 2015). The topology is similar in pattern with those of Murakami et al. (2012a, 2012b, 2014, 2015), in which Pterophocaena nishinoi is reconstructed as the earliest diverging phocoenid; all phocoenids from Hokkaido are placed as a clade; Lomacetus and Australithax as a clade; and extant species as monophyletic. Colpaert et al. (2015) reconstructed Pterophocaena nishinoi basal to all phocoenids as similar to this study, but among an unresolved polytomy with Delphinodon dividum and Kentriodon pernix. Both in our study and the Murakami studies, just crown-ward from Pterophocaena nishinoi, a clade of Archaeophocaena teshioensis and M iophocaena nishinoi are recovered. Colpaert et al. (2015) placed A. teshioensis and Miophocaena nishinoi with Semirostrum ceruttii as an unresolved polytomy. This study shows an unresolved polytomy, which includes Salumiphocaena + P. tedfordi + P. longirostris + (Septemtriocetus + Brabocetus + Semirostrum) + an extant clade. Salumiphocaena stocktoni was reported from the early late Miocene (12.6 to 9.0 Ma) Valmonte Diatomite Member in California (Wilson, 1973; Barnes, 1985a), which is possibly the oldest known record of the Phocoenidae (Uhen et al., 2008). Semirostrum ceruttii is placed as one of the most crownward extinct species in our study and also Racicot et al. (2014). Just basal from the extant clade, our study has a clade of Septemtriocetus + Brabocetus + Semirostrum. In Colpaert et al. (2015), Brabocetus was placed just basal to the Septemtriocetus. The three species of Piscolithax do not form a clade in this study and the previous studies, except Fajardo‐Mellor et al. (2006).
DISCUSSION
Comparison Between the Skulls of Numataphocoena yamashitai Holotype and NFL 2074
Numataphocoena yamashitai can be diagnosed by having the maxillary terrace (new term). The maxillary terrace is a raised area along a sulcus from the medial border of the posterior dorsal infraorbital foramen. Modern phocoenids also have the raised area and sulci from the posterior dorsal infraorbital foramen, but they are much shorter sulcus and weaker raised area than in Numataphocoena yamashitai.
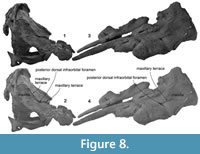 The maxillary terrace is a natural structure, not a taphonomic artifact because the internal wall of the braincase does not show any deformations (holotype; NFL 7 and referred specimen; NFL 2074).
The maxillary terrace is a natural structure, not a taphonomic artifact because the internal wall of the braincase does not show any deformations (holotype; NFL 7 and referred specimen; NFL 2074).
Comparison of the maxillary terrace in the holotype NFL 7 and the referred skull NFL 2074 reveals that the relationships of the maxillary terrace and the dorsal infraorbital foramen are different (Figure 8). On NFL 2074, the maxillary terrace is restricted by a sulcus extending from the medial border of the posterior dorsal infraorbital foramen, and its length is about 46 mm. On the other hand, the holotype NFL 7 shows an extra-extended sulcus, which continues anterior to the posterior dorsal infraorbital foramen. The anterior part of the maxillary terrace is broken in NFL 7, and the length of the sulcus is 55+ mm. The anterior part of the sulcus is seen only on the holotype. Additional skulls are required to consider the maxillary terrace variation.
In addition to the maxillary terrace, some morphological differences exist between NFL 7 and NFL 2074. The nuchal crest is different in the degree of development. In NFL 7 the nuchal crest is well-developed and rises dorsally, and its anterior margin is weakly bowed down anteriorly, continuing to the lateral margin of the frontal boss. On the other hand, NFL 2074 has an incipient crest on the preserved lateral part of the frontal. These differences may be due to ontogenetic variation as NFL 2074 is discussed as younger individual based on the skull size and fusions of the skull sutures. However, in an ontogenetic study of Stenella coeruleoalba, Ito and Miyazaki (1990) reported sexual dimorphism of the skull. Females over three years of age have a developed nuchal crest, compared to males, in which the nuchal crest is not developed at any growth stage. Thus, if Numataphocoena yamashitai had sexual variation on the nuchal crest like Stenella coeruleoalba, and if NFL 2074 was a different sex to NFL 7, NFL 2074 might not have developed a conspicuous nuchal crest even at attainment of physical maturity.
A Clade of Fossil Phocoenids from Hokkaido
The phylogenetic results show a clade of fossil phocoenids, which includes Archaeophocaena, Miophocaena, and Haborophocoena spp. and Numataphocoena from the late Miocene to early Pliocene of Hokkaido, Japan. As an exception, Pterophocaena nishinoi, one of the phocoenids from Hokkaido is not included in the clade of fossil phocoenids from Hokkaido, and is reconstructed as just crown to the outgroup Kentriodon in our analysis. The clade of Hokkaido phocoenids has never been discussed before although the clade appeared in the appendix in an implied analysis (down weighting homoplastic characters) of Tanaka and Fordyce (2015), which is the original matrix of this study. A similar clade is seen in the results of Colpaert et al. (2015), which included four Hokkaido phocoenids (Haborophocoena spp., Miophocaena and Archaeophocaena).
The clade of Hokkaido phocoenids in this study is supported by three synapomorphies; the premaxillary foramen medial to the center of the premaxilla (Character 22), wide premaxillae compared to the rostrum at antorbital notch (Character 34), and the anterior-most dorsal infraorbital foramen lies posterior to the level of the antorbital notch (Character 114). A fossil phocoenid from Hokkaido, Pterophocaena nishinoi has the premaxillary foramen at the midpoint of the premaxilla (Character 22), and the anterior-most dorsal infraorbital foramen is located anterior to the antorbital notch (Character 114). Having a low premaxillary eminence is one of the diagnostic features of the clade of Hokkaido phocoenids. Pterophocaena does not have the premaxillary eminence. The low premaxillary eminence is also seen in the Atlantic species, Brabocetus and Septemtriocetus.
 Our analysis recognizes two subclades, rather than having a pectinate topology of the stem taxa in which the clade of Hokkaido phocoenids (Numataphocoena and Haborophocoena) and another clade (Lomacetus, Piscolothax, Semirostrum and other species + extant species). The clade of Hokkaido phocoenids is branching immediately with a clade of the extant + Eastern Pacific + Atlantic clades. It means that the Phocoenidae was separated into Eastern and Western Pacific clades in the Late Miocene to Early Pliocene, early in the history of phocoenids (Figure 9).
Our analysis recognizes two subclades, rather than having a pectinate topology of the stem taxa in which the clade of Hokkaido phocoenids (Numataphocoena and Haborophocoena) and another clade (Lomacetus, Piscolothax, Semirostrum and other species + extant species). The clade of Hokkaido phocoenids is branching immediately with a clade of the extant + Eastern Pacific + Atlantic clades. It means that the Phocoenidae was separated into Eastern and Western Pacific clades in the Late Miocene to Early Pliocene, early in the history of phocoenids (Figure 9).
Phocoenids from Hokkaido phocoenids includes two subclades, the Archaeophocaena + Miophocaena clade and the Haborophocoena toyoshimai + (Numataphocoena yamashitai + Haborophocoena minutus) clade.
The branching pattern of fossil phocoenids from Hokkaido in previous works required the unnecessarily long ghost lineages in each branch (Murakami et al., 2012a, 2012b, 2014). Now, the phocoenids from Hokkaido as a clade makes the branching hypothesis of phocoenids highly valid based on parsimonious recognition because of minimizing a geological range of ghost linages of each branch (Figure 9). Although the phocoenids from Hokkaido are geologically younger in age, early Pliocene, than those from the Northeastern Pacific and the Atlantic such as the early late Miocene Lomacetus, Australithax and Salumiphocaena, and Piscolithax spp. from the latest Miocene, the former species have been recognized as being earlier diverging. The new branching pattern in this study shows that the common ancestor of the two subgroups appeared before the early late Miocene, based on the topology and the oldest known records of each subgroup. Moreover, our analyses revealed that the fossil phocoenids from Hokkaido was unrelated to the extant clade.
This study uses larger numbers of taxa and characters than Colpaert et al. (2015). The results of Colpaert et al. (2015) and our study show a similar pattern in terms of recognizing two subclades, rather than having a pectinate topology. In the Colpaert et al. (2015), the fossil phocoenids from Hokkaido were in a clade with Semirostrum. But, in our result, Semirostrum is placed among the unresolved polytomy, which is outside of the clade of fossil phocoenids from Hokkaido.
Phylogenetic Relationships and Morphologies of Numataphocoena yamashitai and Haborophocoena spp
Numataphocoena yamashitai and Haborophocoena minutus are located in the clade of phocoenids from Hokkaido, which is supported by two synapomorphies, that is, the relatively large exposure of the lacrimojugal in ventral view (between 62% and 69% of anteroposterior distance from antorbital notch to postorbital ridge, (Character 15) and narrow premaxillae at the level of the antorbital notch (Character 21). Haborophocoena toyoshimai is sister to the clade containing Numataphocoena yamashitai + Haborophocoena minutus. The three species is supported by one synapomorphy: flat (not bent) posterior process of the periotic in lateral view (Character 84).
The genus Haborophocoena is paraphyletic in the result of this study and also previous studies, which included the two species of Haborophocoena (see next). [AUTHOR: what is see next? Next sentence, next paragraph? I can link to it if you identify] Some studies found Haborophocoena minutus into a clade with later branching phocoenids and the extant phocoenids (Murakami et al., 2012a, 2012b, 2014, 2015; Tanaka and Fordyce, 2015). Colpaert et al. (2015) found Haborophocoena minutus in a clade with Archaeophocaena + Miophocaena + Semirostrum clade.
The character state shared only between the two species of Haborophocoena but not with Numataphocoena is having narrow mandibular fossae of the squamosals (Character 112). Numataphocoena yamashitai shows a wider mandibular fossa of the squamosal, which receives the mandibular condyle. This character is changed in the two lineages independently in this analysis, and might be related with mandibular movement and/or feeding styles. The preserved posterior left mandible of Haborophocoena toyoshimai shows a wider and more tilted clockwise mandibular condyle, compared to Phocoena phocoena, which has a narrower mandibular condyle. Numataphocoena yamashitai does not preserve the mandibular condyle. In short, these mandibular variations among the three species suggest that they might have different feeding ecology.
CONCLUSION
The new referred skull (NFL 2074) of Numataphocoena yamashitai from the upper part of the Horokaoshirarika Formation (early Pliocene), Numata, Hokkaido, Japan, adds diagnostic characters of the species, recognizes variations among the species and reveals the phylogenetic position among the Phocoenidae. Our cladistic analysis places Numataphocoena yamashitai adjacent to Haborophocoena toyoshimai and Haborophocoena minutus, among a clade of early branching phocoenids, all of which are chronologically and geographically close to each other, being all from Hokkaido. Numataphocoena yamashitai differs from other phocoenids in having a maxillary terrace, narrower and sharper anterior part of the internal acoustic meatus, and a robust anterior process of the periotic. NFL 2074, which is about 80% size of the holotype, NFL 7, is probably younger than the physically mature holotype. NFL 2074 does not have a well-developed nuchal crest like the holotype skull, which is probably the result of intraspecific variation.
ACKNOWLEDGMENTS
We thank R. Nakashima (National Institute of Advanced Industrial Science and Technology) for deposition of the specimen . We thank T. Shinmura (Ashoro Fossil Museum) for providing the restoration of Numataphocoena yamashitai. We thank R. Racicot (Howard University) and R. Boessenecker (College of Charleston) for giving constructive comments through reviewing, which improved the manuscript.Thanks go to O. Lambert (Institut royal des Sciences naturelles de Belgique), Racicot (HU) and M. Murakami (Shumei University) for giving helpful suggestions for our previous version of this manuscript. Thanks also go to M. Kimura (Numata, Fossil Museum, Hokkaido University of Education) and the Numata Kosei Clinic for encouragement to our research. The first author thanks to R.E. Fordyce (University of Otago) for discussion about the morphological characters, and N. Kohno (National Museum of Nature and Science), S. Shinohara (Numata Fossil Museum) and Murakami (Shumei University) for discussion on geology and chronology of phocoenids. We thank to H. Ito (National Research Institute of Fisheries Science, Fisheries Research Agency) for discussion about sexual dimorphisms on Stenella coeruleoalba, and H. Furusawa (Sapporo Museum Activity Center) for his permission to examine specimens stored in his institution for comparison.
REFERENCES
Aguirre-Fernández, G., Barnes, L.G., Aranda-Manteca, F.J., and Fernández-Rivera, J.R. 2009. Protoglobicephala mexicana, a new genus and species of Pliocene fossil dolphin (Cetacea; Odontoceti; Delphinidae) from the Gulf of California, Mexico. Boletin de la Sociedad Geologica Mexicana, 61:245-265.
Allen, G.M. 1923. The black finless porpoise, Meomeris. Bulletin of the Museum of Comparative Zoology, 65:233-256.
Arnold, P.W. and Heinsohn, G.E. 1996. Phylogenetic status of the Irrawaddy dolphin Orcaella brevirostris (Owen in Gray): a cladistic analysis. Memoirs of the Queensland Museum, 39:141-204.
Barnes, L.G. 1984a. Fossil odontocetes (Mammalia: Cetacea) from the Almejas Formation, Isla Cedros, Mexico. Paleobios, 42:1-46.
Barnes, L.G. 1984b. Whales, dolphins and porpoises; origin and evolution of the Cetacea, p. 139-158. In Gingerich, P.D. and Badgle, C.E. (eds.), Mammals. Notes for a Short Course, University of Tennessee, Department of Geological Science.
Barnes, L.G. 1985a. Evolution, taxonomy and antitropical distributions of the porpoises (Phocoenidae, Mammalia). Marine Mammal Science, 1:149-165.
Barnes, L.G. 1985b. Fossil pontoporiid dolphins (Mammalia: Cetacea) from the Pacific coast of North America. Contributions to Science, Natural History Museum of Los Angeles County, 363:1-34.
Barnes, L.G. 1990. The fossil record and evolutionary relationships of the genus Tursiops, p. 3-26. In Leatherwood, S. and Reeves, R.R. (eds.), The Bottlenose Dolphin. Academic Press Inc., San Diego, New York.
Bianucci, G. 2005. Arimidelphis sorbinii a new small killer whale-like dolphin from the Pliocene of Marecchia River (Central eastern Italy) and a phylogenetic analysis of the Orcininae (Cetacea: Odontoceti). Rivista Italiana di Paleontologia e Stratigrafia, 111:329-344.
Boessenecker, R.W. 2013. A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans. Geodiversitas, 35:815-940.
Brisson, A. 1762. Regnum Animale in Classes IX distributum sive synopsis methodica. Edito altero auctior. Theodorum Haak, Leiden, 296.
Burmeister, G. 1885. Examen crítico de los mamíferos y reptiles fósiles denominados por D. Augusto Bravard y mencionados en su obra precedente, Anales del Museo Nacional de Buenos Aires, p. 95-174.
Colpaert, W., Bosselaers, M., and Lambert, O. 2015. Out of the Pacific: a second fossil porpoise from the Pliocene of the North Sea Basin. Acta Paleontologica Polonica, 60:1-10.
Curry, B.E. 1992. Facial anatomy and potential function of facial structures for sound production in the harbor porpoise (Phocoena phocoena) and Dallʼs porpoise (Phocoenoides dalli). Canadian Journal of Zoology, 70:2103-2114.
de Muizon, C. 1983. Un nouveau Phocoenidae (Cetacea) du Pliocène inférieur du Pérou. Comptes Rendus de l’Académie des Sciences, Paris, Série II, 296:1203-1206.
de Muizon, C. 1984. Les vertébrés fossiles de la Formation Pisco (Pérou). deuxiéme partie: les Odontocétes (Cetacea, Mammalia) du Pliocéne inférieur de Sud-Sacaco. Travaux de l’Institut Français d’Études Andines, 27:1-188.
de Muizon, C. 1985. Nouvelles données sur le diphylétisme des Dauphins de rivière (Odontoceti, Cetacea, Mammalia). Comptes Rendus l'Academie des Sciences series 2, 301:359-362.
de Muizon, C. 1987. The affinities of Notocetus vanbenedeni, an Early Miocene platanistoid (Cetacea, Mammalia) from Patagonia, southern Argentina. American Museum Novitates, 2904:1-27.
de Muizon, C. 1988. Les relations phylogenetiques des Delphinida (Cetacea, Mammalia). Annales de Paléontologie, 74:159-227.
de Muizon, C. 1991. A new Ziphiidae (Cetacea) from the Early Miocene of Washington State (USA) and phylogenetic analysis of the major groups of odontocetes. Bulletin du Muséum National d'Histoire Naturelle, 12:279-326.
de Muizon, C. 1994. Are the squalodonts related to the platanistoids? Proceedings of the San Diego Society of Natural History, 29:135-146.
De Smet, W.M.A. 1977. The regions of the cetacean vertebral column volume 3, p. 59-80. In Harrison, R.J. (ed.), Functional Anatomy of Marine Mammals. Academic Press, London.
Fajardo-Mellor, L., Berta, A., Brownell, R.L., Boy, C.C., and Goodall, N.P. 2006. The phylogenetic relationships and biogeography of true porpoises (Mammalia: Phocoenidae) based on morphological data. Marine Mammal Science, 22:910-932.
Flower, W.H. 1867. Description of the skeleton of Inia geoffrensis and the skull of Pontoporia blainvillii, with remarks on the systematic position of these animals in the Order Cetacea. Transactions of the Zoological Society of London, 6:87-116.
Flower, W.H. 1884. On the characters and divisions of the Family Delphinidae. Proceedings of the Zoological Society of London, 1883:466-513.
Fordyce, R.E. 1989. Origins and evolution of Antarctic marine mammals. Origins and Evolution of the Antarctic Biota, Geological Society Special Publiation, 47:269-281.
Fordyce, R.E. 1994. Waipatia maerewhenua , new genus and new species (Waipatiidae, new family), an archaic Late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand. Proceedings of the San Diego Society of Natural History, 29:147-176.
Fordyce, R.E. 2002. Simocetus rayi (Odontoceti: Simocetidae, new family): A bizarre new archaic Oligocene dolphin from the eastern North Pacific. Smithsonian Contributions to Paleobiology, 93:185-222.
Fordyce, R.E. and de Muizon, C. 2001. Evolutionary history of whales: a review, p. 169-234 In Mazin, J.-M. and de Buffrenil, V. (eds.), Secondary Adaptation of Tetrapods to Life in Water. Pfeil, München, Germany.
Fordyce, R.E., Quilty, P.G., and Daniels, J. 2002. Australodelphis mirus, a bizarre new toothless ziphild-like fossil dolphin (Cetacea : Delphinidae) from the Pliocene of Vestfold Hills, East Antarctica. Antarctic Science, 14:37-54.
Fraser, F.C. and Purves, P.E. 1960. Hearing in cetaceans: evolution of the accessory air sacs and the structure of the outer and middle ear in recent cetaceans. Bulletin of the British Museum of Natural History (Zoology) 7:1-140.
Geisler, J.H., Godfrey, S.J., and Lambert, O. 2012. A new genus and species of late Miocene inioid (Cetacea, Odontoceti) from the Meherrin River, North Carolina, USA. Journal of Vertebrate Paleontology, 32:198-211.
Geisler, J.H., McGowen, M.R., Yang, G., and Gatesy, J. 2011. A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evolutionary Biology, 11:1-33.
Geisler, J.H. and Sanders, A.E. 2003. Morphological evidence for the phylogeny of Cetacea. Journal of Mammalian Evolution, 10:23-129.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics, 24:774-786.
Gray, J.E. 1825. An outline of an attempt at the disposition of Mammalia into tribes and families, with a list of the genera apparently appertaining to each tribe. Philosophical Annals, 26:337-344.
Heyning, J.E. 1989. Comparative facial anatomy of beaked whales (Ziphiidae) and a systematic revision among the families of extant Odontoceti. Contributions in Science, Natural History Museum of Los Angeles County., 405:1-64.
Heyning, J.E. 1997. Sperm whale phylogeny revisited: analysis of the morphological evidence. Marine Mammal Science, 13:596-613.
Ichishima, H. 2011. Do cetaceans have the mesethmoid? Memoir of the Fukui Prefectural Dinosaur Museum, 10:63-75.
Ichishima, H. and Kimura, M. 2000. A new fossil porpoise (Cetacea: Delphinoidea: Phocoenidae) from the Early Pliocene Horokaoshirarika Formation, Hokkaido, Japan. Journal of Vertebrate Paleontology, 20:561-576.
Ichishima, H. and Kimura, M. 2005. Haborophocoena toyoshimai, a new Early Pliocene porpoise (Cetacea; Phocoenidae) from Hokkaido, Japan. Journal of Vertebrate Paleontology, 25:655-664.
Ichishima, H. and Kimura, M. 2009. A new species of Haborophocoena, an Early Pliocene phocoenid cetacean from Hokkaido, Japan. Marine Mammal Science, 25:855-874.
Ichishima, H. and Kimura, M. 2013. New material of Haborophocoena toyoshimai (Odontoceti: Phocoenidae) from the lower Pliocene Embetsu Formation of Hokkaido, Japan. Paleontological Research, 17:127-137.
Ito, H. and Miyazaki, N. 1990. Skeletal development of the striped dolphin (Stenella coeruleoalba) in Japanese waters. Journal of the Mammalogical Society of Japan, 14:79-96.
Kasuya, T. 1973. Systematic consideration of recent toothed whales based on the morphology of tympano-periotic bone. Scientific Reports of the Whales Research Institute, Tokyo, 25:1-103.
Kobayashi, I., Hata, M., Yamaguchi, S., and Kakimi, T. 1969. Geology of the Moseushi district. <Quadrangle series scale 1:50,000, Asahikawa (3). Geological Survey of Japan, Kawasaki, Japan.
Kohno, N., Tomida, Y., Hasegawa, Y., and Furusawa, H. 1995. Pliocene tusked odobenids (Mammalia: Carnivora) in the western North Pacific, and their paleobiogeography. Bulletin-National Science Museum Tokyo Series C, 21:111-130.
Lambert, O. 2005. Phylogenetic affinities of the long-snouted dolphin Eurhinodelphis (Cetacea, Odontoceti) from the Miocene of Antwerp, Belgium. Palaeontology, 48:653-679.
Lambert, O. 2008. A new porpoise (Cetacea, Odontoceti, Phocoenidae) from the Pliocene of the North Sea. Journal of Vertebrate Paleontology, 28:863-872.
Luo, Z. and Marsh, K. 1996. Petrosal (periotic) and inner ear of a Pliocene kogiine whale (Kogiinae, Odontoceti): implications on relationships and hearing evolution of toothed whales. Journal of Vertebrate Paleontology, 16:328-348.
Maddison, W.P. and Maddison, D.R. 2011. Mesquite: a modular system for evolutionary analysis. Available at http://mesquiteproject.org.
Marsh, H., Lloze, R., Heinsohn, G.E., and Kasuya, T. 1989. Irrawaddy dolphin - Orcaella brevirostris, (Gray, 1866), p. 101-118. In Ridgway, S.H. and Harrison, S. (eds.), Handbook of Marine Mammals. Volume 4: River Dolphins and the Larger Toothed Whales.
Mead, J.G. 1975. Anatomy of the external nasal passages and facial complex in the Delphinidae (Mammalia, Cetacea). Smithsonian Contributions to Zoology, 207:1-72.
Mead, J.G. and Fordyce, R.E. 2009. The therian skull: a lexicon with emphasis on the odontocetes. Smithsonian Contributions to Zoology, 627:1-248.
Messenger, S.L. and McGuire, J.A. 1998. Morphology, molecules, and the phylogenetics of cetaceans. Systematic Biology, 47:90-124.
Miller, G.S. 1923. The telescoping of the cetacean skull. Smithsonian Miscellaneous Collections, 76:1-70.
Murakami, M., Shimada, C., Hikida, Y., and Hirano, H. 2012a. A new basal porpoise, Pterophocaena nishinoi (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene of Japan and its phylogenetic relationships. Journal of Vertebrate Paleontology, 32:1157-1171.
Murakami, M., Shimada, C., Hikida, Y., and Hirano, H. 2012b. Two new extinct basal phocoenids (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene Koetoi Formation of Japan and their phylogenetic significance. Journal of Vertebrate Paleontology, 32:1172-1185.
Murakami, M., Shimada, C., Hikida, Y., and Hirano, H. 2015. New fossil remains from the Pliocene Koetoi Formation of northern Japan provide insights into growth rates and the vertebral evolution of porpoises. Acta Palaeontologica Polonica, 60:97-111.
Murakami, M., Shimada, C., Hikida, Y., Soeda, Y., and Hirano, H. 2014. Eodelphis kabatensis, a new name for the oldest true dolphin Stenella kabatensis Horikawa, 1977 (Cetacea, Odontoceti, Delphinidae), from the upper Miocene of Japan, and the phylogeny and paleobiogeography of Delphinoidea. Journal of Vertebrate Paleontology, 34:491-511.
Nakashima, R. and Majima, R. 2000. The nature of shell beds in the inner-shelf deposits-A case study of the upper Miocene to lower Pliocene Horokaoshirarika Formation in central Hokkaido. Journal of Geological Society of Japan (Chishitsugakuzassi), 106:136-150. (In Japanese with English abstract)
Nakashima, R. and Watanabe, M. 2000. First occurrence age of Fortipecten takahashii (Yokoyama) (Bivalvia: Pectinidae) from the lower part of the upper Miocene Horokaoshirarika Formation in Numata-cho, central Hokkaido. The Journal of the Geological Society of Japan, 106:578-581. (In Japanese with English abstract)
Noble, B. and Fraser, F. 1971. Description of a skeleton and supplementary notes on the skull of a rare porpoise Phocoena sinus Norris & McFarland 1958. Journal of Natural History, 5:447-464.
Racicot, R.A., Deméré, T.A., Beatty, B.L., and Boessenecker, R.W. 2014. Unique feeding morphology in a new prognathous extinct porpoise from the Pliocene of California. Current Biology, 24:774-779.
Rommel, S. 1990. Osteology of the bottlenose dolphin, p. 29-49. In Leatherwood, S. and Reeves, R.R. (eds.), The Bottlenose Dolphin . Academic Press Inc, San Diego, New York.
Sanders, A.E. and Barnes, L.G. 2002. Paleontology of the late Oligocene Ashley and Chandler Bridge formations of South Carolina, 2: Micromysticetus rothauseni, a primitive cetotheriid mysticete (Mammalia: Cetacea). Smithsonian Contributions to Paleobiology, 93:271-293.
Tanaka, Y. 2016. A new and ontogenetically younger specimen of Numataphocoena yamashitai from the lower Pliocene, the upper part of the Horokaoshirarika Formation, Numata, Hokkaido, Japan. Paleontological Research, 20:105-115.
Tanaka, Y. and Fordyce, R.E. 2014. Fossil dolphin Otekaikea marplesi (latest Oligocene, New Zealand) expands the morphological and taxonomic diversity of Oligocene cetaceans. PLoS ONE, 9:e107972.
Tanaka, Y. and Fordyce, R.E. 2015. A new Oligo-Miocene dolphin from New Zealand: Otekaikea huata expands diversity of the early Platanistoidea. Palaeontologia Electronica, 18(2.23A):1-71.
Tomida, Y. and Kohno, N. 1992. Fossil marine mammals from the Koetoi Formation (Middle Late Miocene to Early Pliocene) in Wakkanai City, northern Hokkaido, Japan. Memoirs of the National Science Museum (Tokyo), 25:49-56.
Uhen, M.D., Fordyce, R.E., and Barnes, L.G. 2008. Odontoceti, p. 566-606. In Janis, C.M., Gunnell, G.R., and Uhen, M.D. (eds.), Evolution of Tertiary Mammals of North America, 2. Cambridge University Press, Cambridge.
Wada, N., Ganzawa, Y., Sagayama, T., Takahashi, K., Gocho, M., Watanabe, N., and Akiyama, M. 1986. Stratigraphy and age determination of the pliocene in the Rumoi-Fukagawa district, Hokkaido, Japan, The 93th Annual Meeting of the Geological Society of Japan, p. 142,(In Japanese)
Watanabe, M. and Yoshida, F. 1995. Geology of the Ebishima District. Quadrangle series scale 1:50,000, Asahikawa (3). Geological Survey of Japan, Tsukuba, Japan.
Whitmore, F.C. and Sanders, A.E. 1977. Review of the Oligocene Cetacea. Systematic Zoology, 25:304-320.
Wilson, L.E. 1973. A delphinid (Mammalia, Cetacea) from the Miocene of Palos Verdes Hills, California. University of California Publications in Geological Sciences, 103:1-34.
Yamashita, S. and Kimura, M. 1990. Occurrence of Early Pliocene otariid fossil in Numata-cho, Hokkaido. Earth Science (Chikyu Kagaku) 44:53-60.(In Japanese with English abstract)
